Progenitör hücreden T hücre gelişimi
Hemopoietik kök hücreleri (HKH) → "Thymic seeding progenitor" (TSP) hücreleri → "early thymic progenitor" (ETP) hücreleri → DN1 → DN2 → DN3 → DN4 → CD4 veya CD8.
- CD4 ve CD 8 "Double-negative" (DN) timositler
- CD4 ve CD8 "Double-positive" (DP) timositler
- CD4 veya CD8 moleküllerinden birini içeren "Single-positive" T hücreleri
Self peptidlerden MHC klas I kompleksine bağlanan TCR'ye sahip T hücreleri CD8+ ve self peptidlerden MHC klas II kompleksine bağlananlar ise CD4+ T hücreleri adın alır. Bu hücreler artık medulladan perifer lenf bölgelerine gitmeye hazırdır (Single positive - SP).
Başarılı bir şekilde pre-TCR eksprese etmeyi başaranlar double positive (DP) olur ve TCR-α zincirini eksprese eden gen lokusunda, genlerin tekrar düzenlenmesi ile pre-TCR-α zincirinin yerini tekrar düzenlenmiş TCR α-zinciri alır. DP hücrelerde, CD4 veya CD8 ayırımı yapmadan αβ TCR tamamlanır.
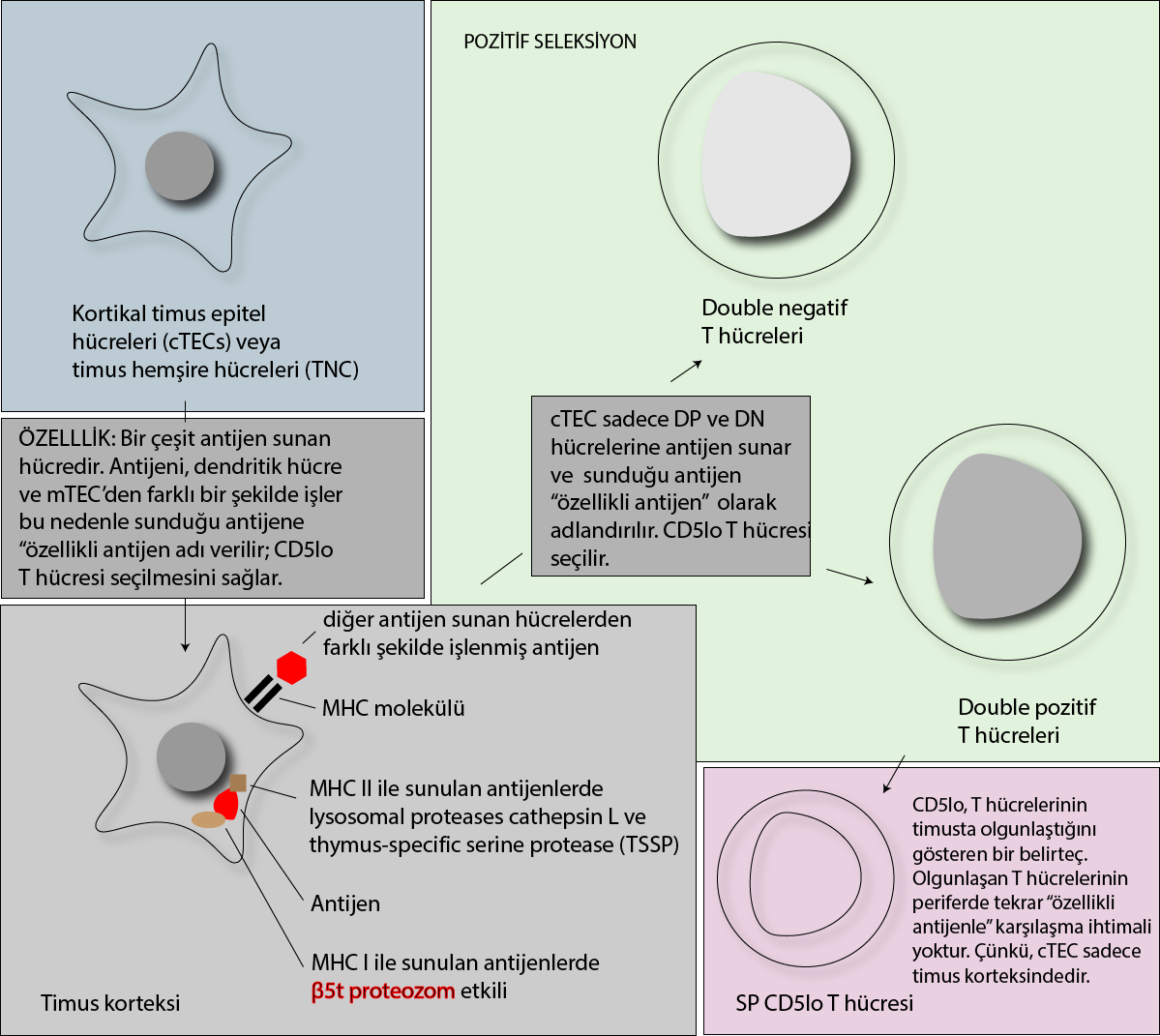
αβ TCR+CD4+CD8+ (DP) timositler, self ile ilgili yüksek dansitede MHC klas I ve II moleküllerini eksprese eden kortikal epitel hücreleri ile etkileşime girer.
Santral tölerans
DP pozitif timositlerin kaderi TCR ile self peptid MHC ligandı arasındaki etkileşim sonucu belirlenir; bağlanma sonucu oluşan sinyal çok zayıf ise yaşmaya devam etmelerini sağlayan sinyali oluşmadığı için bir süre sonra ölürler (bir nevi gecikmiş apopitoz denilebilir). Kortekste, uygun bir şekilde orta düzeyde TCR sinyali olan hücrelerde effektif matürasyon başlatılır (pozitif seleksiyon).
Timositler daha sonra medullaya göç eder. Medullada timositler negatif seleksiyona uğrarlar. Bu aşamada, antijen sunan hücreler (dendritik hücre veya makrofaj gibi) timostilere self antijen sunarlar. Self antijenle güçlü bir şekilde bağlanan TCR, çok fazla sinyal oluşmasına neden olur ve akut apopitoz başlar (negatif seleksiyon).
CD4 ve CD 8 "Double-negative" (DN) timositler
TSP hücreleri, timusta ETP hücrelerine dönüşür. ETP hücreleri, heterojen DN1 hücrelerinin alt grubudur ve dış kortekse doğru ilerlerken 4 basamakta farklılaşır (DN1, CD44+CD25-; DN2, CD44+CD25+; DN3, CD44-CD25+; and DN4, CD44-CD25-). Alt grup sınıflaması, hücrenin CD44 (bir adezyon molekülüdür) ve CD25 (interlökin-2 reseptörü α zinciri) moleküllerinin eksprese edebilme özelliğine göre yapılmaktadır. CD44 eksprese edemeyen fakat CD25 eksprese eden, yani DN3 hücreleri β seleksiyona tabi tutulabilme özelliği kazanır. β seleksiyonda, β zincir genlerinde tekrar düzenleme (rearrangement) yapılır, başarılı bir şekilde TCR-β zinciri eksprese edenler seçilir. β zincirinin başarılı olması durumunda pre-Tα suret zinciri ile birleşebilme özelliğine sahip olması anlamına gelir çünkü bu sayede CD3 ile komplkes oluştrabilecek bir pre-TCR oluşması sağlanır.
- DN1; erken timosit progenitör hücreleri, potansiyel olarak dendritik hücre, NK hücreleri ve makrofajlara dönüşebildiği gibi DN2a hücrelerine de dönüşebilir.
- DN2; DN2a potansiyel olarak dendritik hücre, NK hücreleri ve makrofajlara dönüşebildiği gibi DN2b hücrelerine de dönüşebilir. DN2b γδ T hücrelerine veya DN3a hücrelerine dönüşebilir.
- DN3; DN3a, γδ T hücrelerine veya DN3b hücrelerine dönüşebilir. DN3b ise DN4 hücrelerine dönüşür.
- DN4; double pozitif (DP) hücrelerine dönüşür, TCRα tekrar düzenlemesi yapıldır.
γδ veya αβ zincirler çiftleri
DN'ler γδ T hücreleri veya αβ T hücrelerinden birine dönüşebilir. TCR’de yer alan γ ve β zincir genleri az veya çok eş zamanlı olarak düzenlenmeye (rearrange) başlar. Bu aşamada γ zincir genlerinin düzenlenmesi başarılı olursa o zaman δ zincir genleri de düzenlenmeye başlar. Eğer hem γ hem δ gen düzenlemesi işlevsel ise düzenlenme (rearrangement) işleminin devam etmesine gerek yoktur ve T hücresi γδ T hücresi olarak kalır. Eğer γ ve/veya δ düzenlenmesi işlevsel değilse o zaman β genlerinin düzenlenme işlemi ardından da α genlerinin düzenlenmesi devam eder. Bu durumda αβ ürünler görülmeye başlar ve T hücresi αβ T hücresi olur. Gen düzenleme başarılı olduğunda düzenleme işleminin durmasına allelic exclusion adı verilir. Gen düzenlenmesi başarılı değilse karşılığı olan diğer kromozomda bulunan ikinci gen kopyalarında düzenlenme başlar. Bu girişim de başarısız olursa, hücre apopitoza sevk edilir. T hücre respetörlerinde, B hücrelerindeki gibi somatik hipermutasyon olmaz.
αβ T hücrelerine dönüşenler, DN2'den DN4'e doğru ilerlediğinde, pre-TCR (α-düzenlenmemiş, β-düzenlenmiş) eksprese etmeye başlar. Yani TCR-β zincirini eksprese eden gen lokusunda, genlerin tekrar düzenlenmesi (beta seleksiyon).
- αβ T hücreleri, DN3 basamağında ilk önce düzenlenme yapmayan (non-rearranging) gen lokusundan TCR-α eksprese eder. Pre-T-α, somatik rekombinasyonla tekrar düzenleme (rearrangerment) sonucu oluşan TCR β-zinciri ile çiftleşir. Hücrenin yüzeynideki pre-TCR-αβ çift zinciri sinyal iletimi sağlayan protein grubu (CD3) ile ilişkili olarak belirmektedir. T hücresinin olgunlaşmaya devam etmesi için aktif sinyalleşmeye ihtiyacı vardır.
"Double negative 1" (DN1) hücreler (CD44+CD25-)
DN1 grubunda "early thymic progenitor" (ETP) hücreler yer alır. Bu hücreler yüksek seviyede CD117 eksprese eder. Timustaki hücre havuzunun yaklaşık %0,01 kadarı DN1 hücrelerinden oluşmaktadır. DN1, timusta kortikomedüller bağlantı yerinde yaklaşık 10 gün kadar bulunurlar.
ETP hücrelerinin T hücrelerine doğru ilerlemesi sağlanırken, diğer hücrelere dönüşmesi engellenir. Bunun için Notch sinyali gereklidir. Notch sinyali sayesinde, ETP'nin sadece T hücresine doğru ilerlemesi sağlanır. Notch1 sinyalinde DLL4 (delta-like 4) ligandı rol oynamaktadır. Timus epitel hücrelerindeki DLL4 ligandı silindiğinde T hücrelerin gelişmesi bloke edilir. DLL4 ligand ve Notch1 reseptörü çifti birlikte gereklidir.
"Double negative 2" (DN2) hücreler (CD44+CD25+)
DN1 kortikomedüller bağlantı yerini terk ettikten sonra, timus eptiel hücreleri ve fibroblastlardan aldıkları uyarı sinyalleri doğrultusunda, korteksin derinlerine subkapsüler alana doğru göç eder ve burada DN2 timositlere dönüşür. DN2 aşamasında, hücrenin alternatif hücrelere dönüşmesi biraz daha engellenirken, TCRγ, TCRδ ve TCRβ genlerinde tekrar düzenleme (rearrangement) işlemi başlatılır fakat bu işlem daha sonra DN3 aşamasında bitirilir.
DN2a hücreleri potansiyel olarak dendritik hücrelere ve NK hücrelere dönüşme özelliğinde sahipken DN2b'de sadece NK hücrelerine dönüşebilme potansiyeli devam ederken, dendritik hücrelerine dönüşme potansiyeli kaybolur.
"Double negative 3" (DN3) hücreler (CD44-CD25+)
DN3 aşamasında, hücrenin αβ veya γδ T hücresi tarafında gideceği belirlenir. DN3'te DNA'nın β, γ ve δ lokuslarında fonksiyonel bir TCR zinciri ortaya çıkarmak için yoğun bir şekilde tekrar düzenlemeler devam eder. αβ T hücreleri DN3a'dan DN3b'ye doğru giderken gelişmeye devam eder, bu aşamada β seleksiyon işlemi önem kazanır. Pre-TCR'nin doğru bir şekilde çalışması için, TCR-β'nın doğru bir şekilde düzenlenmiş olması, CD3 zinciri ve invarinat pTα zinciri gerekir. Buna ek olarak en az iki sinyal daha β sekelsiyona katkıda bulunur: kemokin CXCL12'nin reseptörü olan CXCR4 ve Notch1 reseptörü. Bu aşamada Notch1 sinyali hücrenin yaşamaya devam etmesini sağlar.
"Double negative 4" (DN4) hücre (CD44-CD25-)
DN3 aşaması geçildikten sonra, T hücresi olmanın dışında bütün olasılıklar kaybedilir. DN4 aşamasına geçen timosit subkapsüler alanı döner ve tekrar medullaya doğru göç etmeye başlar. Yoldayken, dış kortekste timositler pre-TCR sinyalinden sonra CD4 ve CD8 üretmeye başlar ve öncü hücre olarak timusa geldikten yaklaşık 19 gün sonra "double positive" (DP, CD4+CD8+) aşamasına geçmiş olurlar. Bu aşamada, Rag genleri tekrar eksprese edilmeye başlar ve TCRα rekombinasyonu başlar. T hücrelerinden αβ TCR'si, MHC ligandıyla doğru bir şekilde bağlananların yaşamasında izin verilir ve farklılaşır. Self peptid MHC kompleksiyle orta aviditede etkileşen DP timositler pozitif seleksiyonla ayrılırlar. Yaşayan timositler MHC ile sınırlı TCR eksprese eder. Bundan sonra DP timosit CD4 "single positive" veya CD8 "single positive" hücrelerinden birine doğru gelişir. SP'ler artık medullaya doğru göç eder ve burada negatif seleksiyonla self antijene yüksek afinite ile bağlanan TCR'ye sahip hücreler yok edilir. Bu hücreler sphingosine-1 phospahte receptor 1 (S1P1) üreterek timustan dolaşıma geçer.