B hücrelerinin antijen tarafından uyarılması
B hücrelerine antijen sunulması karmaşıktır, anladığımız kadarıyla bu olay B hücre foliküllerinin perifer ve santralinde meydana gelmektedir. B hücreleri başlangıçta periferde aktive olurlar, genellikle bu olay antijenin makrofaj veya dendritik hücre tarafından sunulmasıyla gerçekleşmektedir. Lenf düğümündeki dendritik hücreler antijen havuzunun ve etkili bağlanmanın gerçekleştiği yere doğru yüzerler. Erkenden gerçekleşen bu olay germinal merkezden ve B hücrelerinindeki BCR afinite olayından bağımsızdır. Geç gerçekleşen olaylarda antijenler germinal merkezde zaten aktive olmuş olan B hücrelerin sunulur, burada FDC antijen sunulmasında merkez rol oynar. FDC'ler uzun süre antijeni tutabilme özelliğine sahiptir bunun yanında günümüzde artık FDC'nin antijenleri yüzeye çıkarma döngüsü nedeniyle hümoral bağışıklığın düzenlenmesinde de aktif rol oynadığı da bilinmektedir. FDC'de bulunan PRR'lerden TLR'ler bu hücrelerin yabancı ve self antijenlere yanıta başka bir seviyede de müdahil olduğunu gösterir.
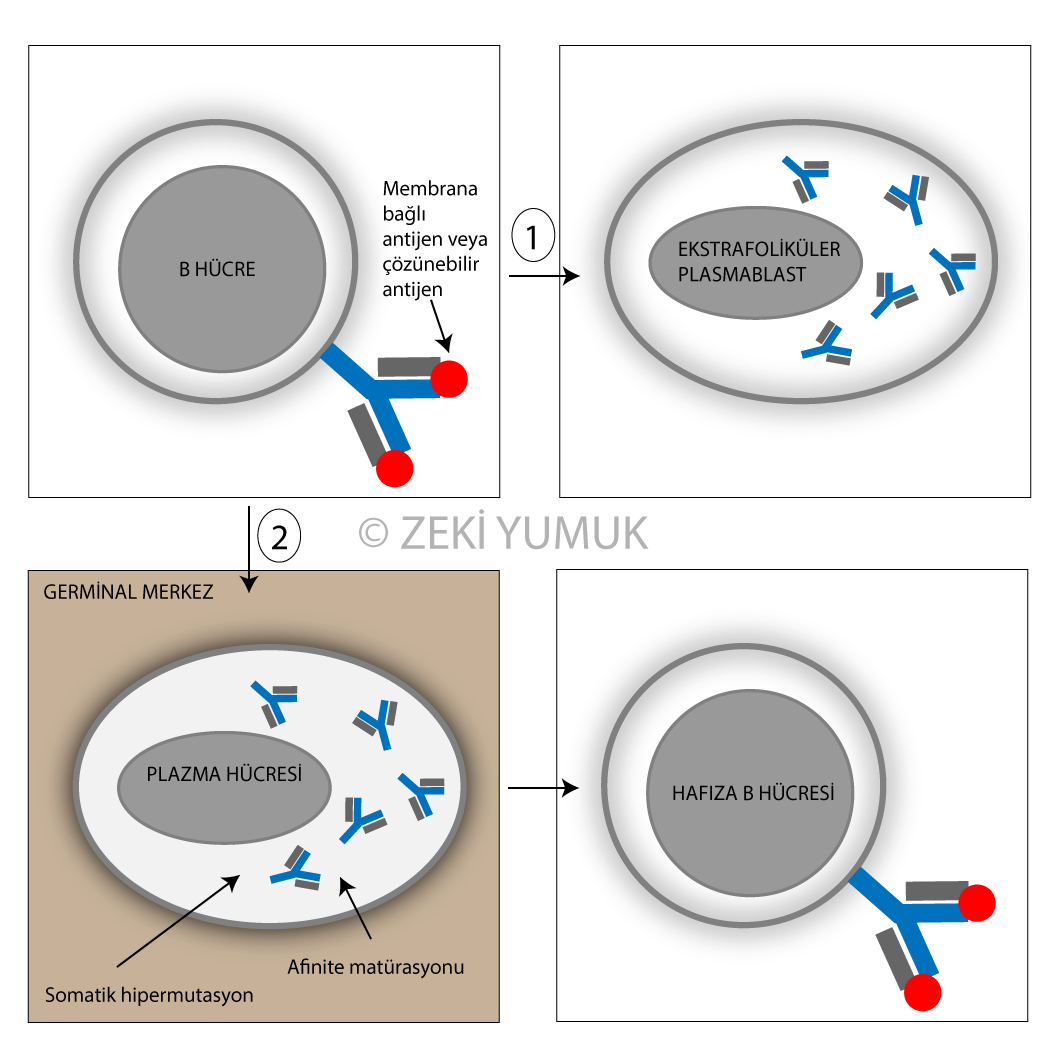
B hücreleri 2 yoldan farklılaşmaktadır. Bir tanesinde, B hücreleri erken dönemde koruyucu yanıt oluşturabilmek için hızlı bir şekilde antikor üreten ektrafoliküler plazmablastlara dönüşürler. İkincisinde ise aktive B hücreleri germinal merkeze girerek afinite matürasyonunu takiben yüksek afiniteli antikor üreten plazma hücrelerine veya tekrar antijenle karşılaşıldığında koruyuculuk sağlayan hafıza B hücrelerine dönüşürler. Antijenlerin çok çeşitleri vardır ve bu nedenle antijene spesifik lenfositlerle antijenlerin karşılaşmalarının hızlı bir şekilde sağlanması gerekir. Antijen çeşidiyle uygun lenfositin karşılaşma olasılığının arttırılması için lenf düğümü ve dalak gibi sekoder lenf organları vardır. Sekonder lenf organlarının yapısı antijen çeşidine uygun lenfositin bulunması ve immün yanıtın başlatılması için uygun bir yapıdır.
Lenfositlerin antijenle karşılaşması mekanizmasının anlaşılması için lenfositlerin lenf organlarında dinlenme fazındaki yerleri ve dinamiklerini anlamak önemlidir. Lenf düğümünde dinlenme fazındaki lenfositler çok hareketlidir. Ancak sadece kendi bölgeleri içerisinde hareket ederler. Bu özellik lenfositlerin sürekli olarak antijen aradığını göstermektedir.
B hücrelerine antijen sunulması
Bağışıklamadan sonra B hücreleri antijeni bulur ve yanıt verir. Küçük molekül ağırlığa sahip çözünür antijenler lenf dokusuna difüzyonlar girdikten sonra karşılaştıkları makrofaj, dendritik hücre ve foliküler dendritik hücre tarafından sunulurlar. Aslında B hücreleri küçük molekül antijenlerle difüzyon aracılığı ile de karşılaşabilir ve sonra folikül ile parakorteks arasındaki sınıra doğru göç eder. Antijenler lenf düğümüne aferent lenf damarları ile getirilir. Subkütan verilen antijenler kısa süre sonra subkapsüler sinüste tespit edilir. B hücrelerin çoğu folikülde bulunduğundan lenf kaynaklı antijenler kanal sisteminden (genellikle parakortekstedir) bağımsız olarak foliküldeki B hücreleri ile karşılaşırlar.
Büyük moleküllerin foliküldeki B hücreleri ile karşılaşması küçük moleküllü antijenlere göre pek mümkün değildir. Bu nedenle bazı özel hücrelerin özellikle subkapsüler sinüste yerleşen hücrelerin büyük antijenleri foliküldeki B hücrelerine taşıması gerekir. Bu hücre makrofajlar olabilmektedir. Makrofajlar B hücreleri subkapsüler alanda etkileşime girdiğinde antijene spesifik B hücresi antijeni, T hücre yardımı için B-T hücre sınırına gitmeden önce BCR ile içeri alır. B hücreleri HEV'den içeri girdiğinde kendilerine antijen sunulması için parakorteks bölgesi idealdir. Nitekim bu bölgede bulunan dendritik hücreler aldıkları antijeni B hücrelerine sunmaktadır. Bu bölgenin diğer bir özelliği ise T hücre yardımının sağlanması için uygun olmasıdır. Dendritik hücrelerin B hücrelerine antijen sunması ile B hücrelerinin aktive olması erkenden o antijene karşı antikor oluşmasını sağlar. Diğer taraftan CD4+ T hücrelerinin aktivasyonundan sonra B hücreleri foliküle göç eder ve burada germinal merkez oluşur. Dalakta dendritik hücrelerin marjinal bölgedeki B hücrelerine antijen sunması sonucu T hücrelerinden bağımsız bir şekilde IgM karekterinde antikor hızlı bir şekilde oluşturduğu görülmektedir.
Klasik olarak FDC'ler sekonder lenf organlarında B hücrelerine antijen sunan hücrelerdir. FDC'ler antijen antikor kompleksindeki antijeni yakalar. Bunu işlem için iki farklı mekanizma vardır: birincisi kompleman sisteme bağlı mekanizmadır. FDC yüksek seviyede kompleman reseptör 1 ve 2 eksprese eder, bu reseptörler C3'ün çeşitli parçalarına bağlanabilmektedir. İkinci mekanizmada ise IgG içeren immün kompleksiler Fc reseptörü ile bağlanır. Her iki mekanizmada da FDC'nin yakaladığı antijen sonucunda immün yanıt başlatılır. Önceden oluşan immünkompleks ve antijenle kapli C3d parçaları B hücrelerin aktivasyonuyla ilişkilidir. Özellikle peritonda bulunan B-1 hücreleri IgM antikorları yapar ve bu IgM antikorları antijenlele oluşturduğu immünkompleksler daha sonra FDC'lerin yüzeyinde birikir. FDC'ler sekonder lenf organlarında antijen depoları olarak görev alabilmektedir, bu sayede vücutta antijen azaldığında bile FDC'ler bulundurmaya devam eder. Bu da, naive B hücresi sekonder lenf organlarında antijenle karşılaştığında düşük afinitede bir etkileşim olması nedeniyle uzun süren etkili bir immün yanıt için gerekmektedir. Bu koşullarda aktive olan B hücresi germinal merkeze gider ve afinite matürasyonu burada başlar.
Germinal merkezde açık ve koyu alanlar vardır. Koyu alanlarda B hücreleri hızlı bir şekilde prolifere olurken açık alanlarda somatik hipermutasyonla yüksek afiniteye sahip BCR eksprese eden B hücre klonları seçilmektedir
Kaynaklar
Batista FD, Harwood NE. The who, how and where of antigen presentation to B cells. Nat Rev Immunol. 2009 Jan;9(1):15-27. doi: 10.1038/nri2454. PMID: 19079135.